De nombreux animaux communiquent par l’intermédiaire de vibrations : les punaises vertes (Nezara virudula) communiquent à l’aide de vibrations transmises par les feuilles sur lesquelles elles se trouvent lors de leur parade amoureuse. De même, les éléphants émettent des signaux vibratoires en frappant le sol de leurs pattes pouvant être reçus par d’autres éléphants à plusieurs kilomètres de distances, selon une étude de O’Connell-Rodwell. Par ailleurs, les araignées communiquent par des vibrations transmises dans la toile et celle ci présente de nombreux aspects : elle est utilisée non seulement lors de la parade amoureuse, mais aussi au sein de sociétés d’araignées sociales et permet la détection d’intrus ou de proie. De plus, le substrat, c’est-à-dire la base matérielle, le support de la communication vibratoire chez les araignées est tout à fait particulier : ce sont les fils de soie, produits par les araignées.
Les araignées ont à leur disposition différents types de signaux pour communiquer. Tout d’abord, des signaux visuels, notamment lors de la parade amoureuse même si elles ont une très mauvaises vue, mais aussi des signaux sonores, en effet les araignées stridulent en frottant différentes parties de leur corps, des signaux chimiques, en effet comme toutes les espèces animales, les araignées communiquent par l’intermédiaire de phéromones, mais c’est bien la communication vibratoire qui est prédominante chez les araignées.
Toutefois, l’étude de la communication vibratoire est très récente, c’est Peter N. Witt, en 1982 qui le premier l’étudia en profondeur dans son ouvrage Spider Communication (voir 14 dans la Bibligraphie) dans lequel il s’intéresse à la communication entre araignées et à la détection des proies, suivant la définition large de la communication comme tout comportement qui transmet une information d’un individu à un autre.
Nous nous intéresserons dans cette partie à la communication vibratoire chez les araignées, tout d’abord en étudiant son rôle, puis comment le signal et produit par l’araignée et transmis dans le substrat avant d’être reçu par une araignée. Enfin nous aborderons les utilisations possibles de cette connaissance par les humains.
Rôle de la communication vibratoire chez les araignées
La communication vibratoire permet principalement aux araignées de repérer des proies et des partenaires sexuels. On peut ensuite définir deux types d’araignées : les araignées sociales et les araignées solitaires. Les araignées solitaires n’ont recours à la communication avec d’autres araignées que lorsqu’elles rencontrent une toile, alors que pour les espèces sociales, soit environ douze genres d’araignées, c’est-à-dire douze regroupements d’espèces, la communication vibratoire a un rôle majeur : elle leur permet d’organiser leur vie en société, et elle leur permet notamment d’organiser la capture des proies en groupe.
De plus, toutes les espèces sociales sont fileuses, elles organisent leur vie dans des structures soyeuses atteignant souvent un volume de 100m3 (voir Figure 1). Ces structures soyeuses, qu’elles ne quittent jamais, favorisent la communication vibratoire. Tout comme pour les araignées solitaires, les vibrations dans la toile leur permettent d’être alertées de la présence d’intrus et de proies : dès qu’un insecte est pris au piège dans la toile, une horde d’araignées l’encercle, le tue puis s’en nourrit. Dès lors, on peut en déduire que la communication vibratoire permet à l’araignée de distinguer ses congénères des proies, et lui permet aussi de localiser la proie sur la toile.

Figure 1 : Toile d’une société d’Anolesimius Eximius, espèce d’araignée sociale (photographie de B. Krafft
Les araignées utilisent aussi les vibrations dans la toile d’une manière qui rappelle l’écholocation chez les chauves-souris ou les dauphins : lorsqu’une proie se tient immobile sur la toile, l’araignée peut envoyer une secousse dans la toile qui provoquera le balancement de la proie, et permettra sa localisation.
Mais certaines araignées utilisent aussi à leur profit la capacité d’autres insectes à communiquer. Par exemple, les sauterelles communiquent normalement par sons qui se propagent dans l’air, mais elles peuvent aussi communiquer par vibrations dans les plantes pour éviter d’attirer les chauves-souris, ces vibrations sont utilisées par les araignées pour localiser les sauterelles, et les chasser. De même, l’araignée sauteuse commune, Portia fimbriata, est capable d’imiter les signaux vibratoires de mâles d’autres espèces d’araignée afin d’attirer les femelles, qu’elle chasse.
Production des signaux vibratoires
Les araignées ont recours à différentes méthodes pour produire des vibrations dans un substrat : les percussions, les vibrations du corps, les stridulations et en tirant directement sur la toile.
La percussion
En frappant le sol de son abdomen ou de son pédipalpe (voir Figure 2), l’araignée peut créer des vibrations qui se propagent dans le sol. La force du signal vibratoire est directement lié à la masse de l’animal à l’origine de la vibration, plus l’animal est massif et plus le rayon de propagation est large.
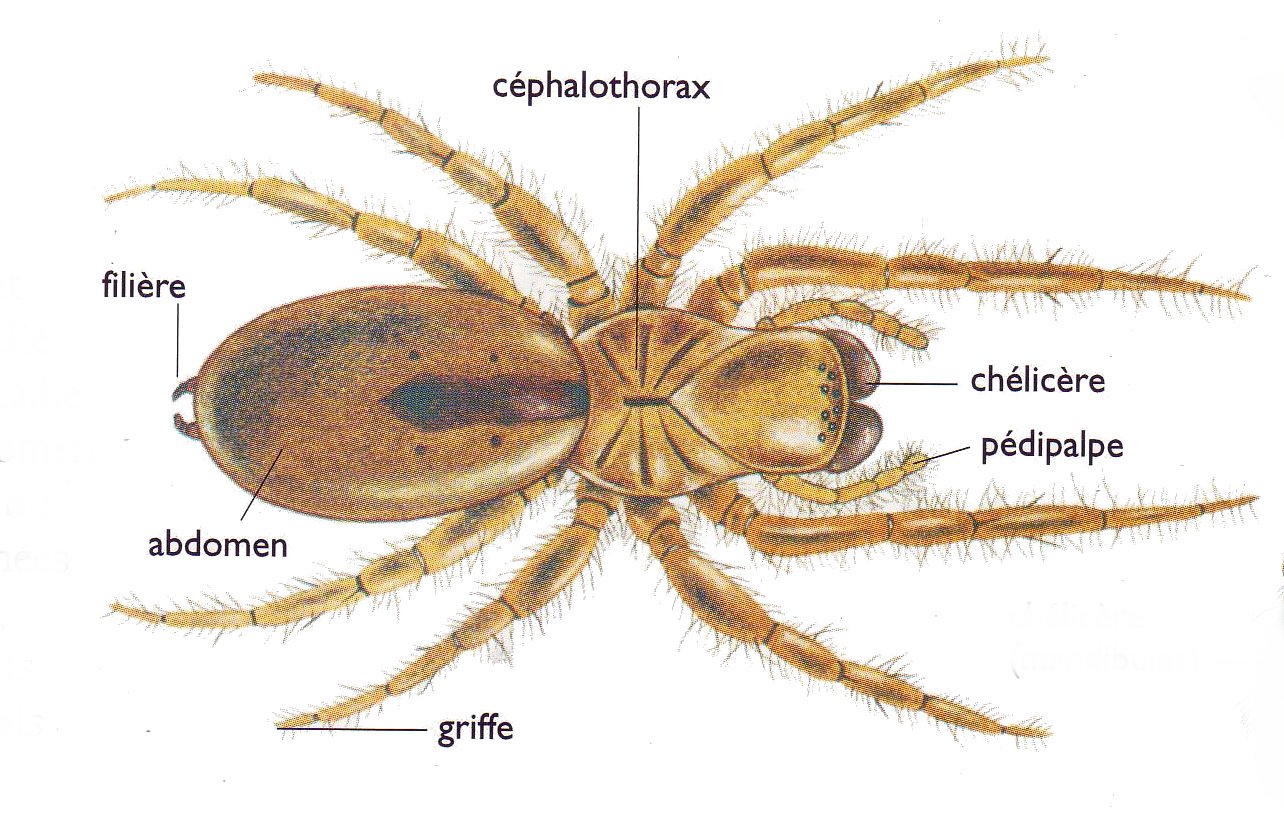
Figure 2 : Anatomie d’une araignée
Ce type d’émission est utilisé surtout par les espèces qui ne font pas de toile, comme les araignées sauteuses (Salticidae) ou les araignées-loups (Lycosidae). Ainsi, le mâle Lycosa gulosa produit des sons qui pourront être perçus par la femelle en frappant le sol de ses pédipalpes.
La trémulation
Les araignées peuvent faire vibrer leur corps afin d’émettre des vibrations qui passeront à travers leurs pattes puis dans le sol, c’est ce qu’on appelle la trémulation. La trémulation ne nécessite pas une morphologie particulière, mais la fréquence des vibrations produites est souvent très faible, car elle est limitée par la vitesse à laquelle l’animal peut faire vibrer son corps, dès lors, elle est donc difficilement analysable.
La stridulation
Les araignées, comme certains insectes, peuvent striduler, c’est-à-dire produire des vibrations en frottant deux parties de leur corps. Ces vibrations sont ensuite transmises au sol par les pattes, et dans l’air sous la forme de sons audibles. Une araignée peut striduler en frottant son abdomen et son céphalothorax, ou en frottant différents appendices, comme patte contre patte, patte contre pédipalpe ou chélicère contre chélicère (voir Figure 2).
Le tiraillement
Les araignées peuvent aussi tirer sur les fils de la toile à l’aide de leur pattes pour y transmettre des vibrations, par exemple en exerçant une forte traction sur la toile puis en relâchant rapidement cette tension, envoyant un signal de grande amplitude.
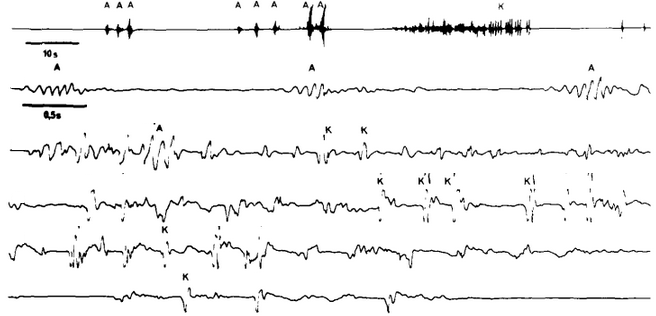
Figure 3 : Vibrations émises par un mâle Tegenaria parietina dans la toile. A : trémulations abdominales ; K : percussions avec l'abdomen (d'après Leborgne et Krafft, 1979). Le signal en ligne 1 est ensuite reproduit avec avec une plus petite échelle aux lignes 2 à 6.
Dans tous ces cas, afin que les signaux soient reconnaissables, les araignées produisent des vibrations caractéristiques en faisant varier la fréquence et l’amplitude des vibrations, ainsi que la répartition de ces vibrations au cours du temps. Nous pouvons voir cette variation du motif des vibrations en Figure 3, on peut remarquer la répétition des trois trémulations abdominales.
Transmission du signal vibratoire
Une fois produit par l’araignée, la vibration se propage dans le substrat, la plupart du temps de la soie d’araignée. C’est pourquoi nous avons effectué différentes expériences pour étudier les propriétés de la soie d’araignée.
Analyses microscopiques
Nous avons procédé à une analyse microscopique de fil de soie d’araignée. Afin d’avoir les fils, nous avons capturé une araignée qui a ensuite fait sa toile dans une boite. Grâce à un appareil photo numérique pour microscope, nous avons pu obtenir des clichés du fil d’araignée.
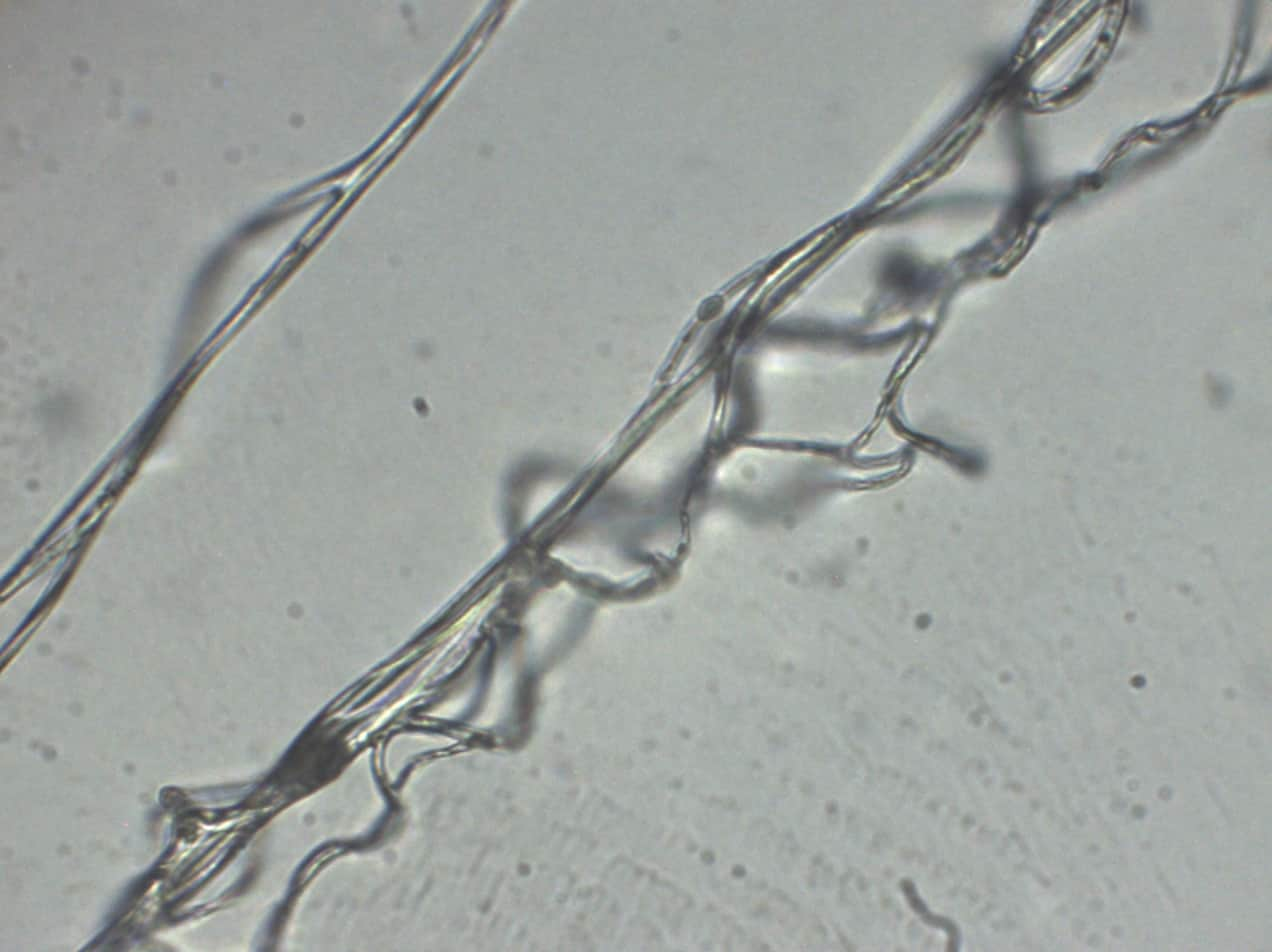
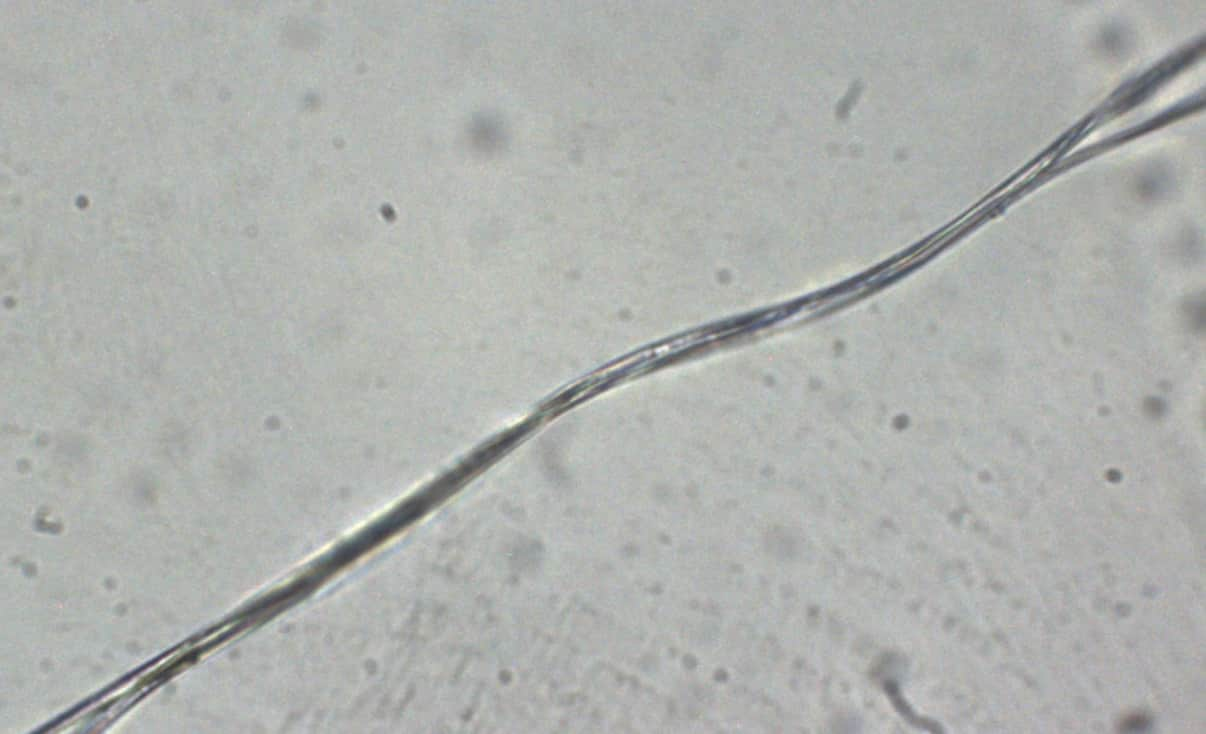
Figure 4 : Photographies de fils d’araignée vus au microscope optique (×600)
Le premier fil n’a pas été coloré, et le second a été coloré au bleu de méthylène. Nous pouvons voir sur ces photographies que les fils d’araignée sont en fait constitués de plusieurs fils tressés les uns autour des autres, toutefois la coloration au bleu de méthylène ne permet pas de différencier différente partie dans chacun des fils.
Mesure de la taille d’un fil de soie d’araignée
Afin d’étudier plus précisément le fil d’araignée, nous avons souhaité mesurer son diamètre. Ceci est possible en analysant le phénomène de diffraction ayant lieu lorsqu’un laser rencontre ce fil d’araignée, c’est-à-dire la manière dont la lumière est diffusée après avoir rencontré le fil.
Lorsqu’on projette la lumière après le fil sur un écran, on obtient des tâches, et la taille de la tâche centrale est inversement proportionnelle au diamètre du fil (ce qu’on appelle « taille de la tâche centrale » correspond en fait à la distance entre le milieu de l’espace entre la tâche centrale et la tâche à sa gauche et le milieu de l’espace entre la tâche centrale et la tâche à sa gauche).

C’est pourquoi à l’aide d’un montage composé d’une diode laser, d’un écran et d’un fil dans une diapositive positionnée entre la diode et l’écran, à un mètre de l’écran, on peut former les tâches sur l’écran et mesurer la taille de la tâche centrale, comme vu en Figure 5.

Figure 5 : Montage pour mesurer la taille du fil de soie
Afin de pouvoir calculer le coefficient de proportionnalité entre le diamètre du fil et l’inverse de la largeur de la tâche centrale, on mesure la taille de la tâche centrale pour des fils de diamètre connu. Par ailleurs on mesure la taille de la tâche centrale d’une lumière de laser diffracté par un fil d’araignée A, puis d’un fil B, prélevé sur la toile de l’araignée capturée. Avec ces mesures, on obtient le tableau suivant.
| Diamètre fil (mm) | Largeur L de la tâche centrale (mm) | 1 / L |
|---|---|---|
| D(filA) | 1,08×102 | 9,26×10-3 |
| D(filB) | 8,0×101 | 1,25×10-2 |
| 4,0×10-2 | 3,3×101 | 3,0×10-2 |
| 6,0×10-2 | 2,2×101 | 4,6×10-2 |
| 1,0×10-1 | 1,3×101 | 7,7×10-2 |
| 1,2×10-1 | 1,1×101 | 9,1×10-2 |
Ces mesures permettent de tracer un graphique. La proportionnalité entre le diamètre du fil et la taille de la tâche centrale est modélisée par une droite de coefficient directeur 0,76 et passant par l’origine du repère.

Figure 6 : 1 / L en fonction du diamètre du fil (L étant la taille de la tâche centrale)
Dès lors on peut calculer :
$[ D(fil) = \frac{\frac{1}{L}}{0.76} $]
Donc :
$[ D(filA) = \frac{9,26\times10^{-3}}{0.76} $]
Soit :
$[ D(filA) = 1,2\times10^{-2} \text{mm} = 1,2\times10^{-5} \text{m} = 12 \mu\text{m} $]
Et,
$[ D(filB) = \frac{1,25\times10^{-2}}{0.76} $]
Soit :
$[ D(filB) = 1,6\times10^{-2} \text{mm} = 1,6\times10^{-5} \text{m} = 16 \mu\text{m} $]
D’après nos mesures, le fil de soie d’araignée à un diamètre ordre 15µm soit environ un dixième de la taille d’un cheveu.
Ainsi, même si le fil de soie d’araignée est extrêmement fin, les araignées sont capables de construire des toiles résistantes au poids de l’araignée elle-même, à celui d’éventuels intrus et à la transmission d’ondes, notamment dans le cadre de la communication.
Vitesse d’une onde dans la toile
Les vibrations émises par l’araignée prennent la forme d’ondes mécaniques progressives dans la toile, c’est-à-dire la propagation d’une perturbation dans un milieu matériel sans transport de matière. Cette onde est dite à une dimension puisqu’elle ne se propage que dans une seule direction.
Afin d’étudier la vitesse d’une onde dans la toile, nous avons décidé de nous intéresser à la vitesse d’une onde dans une corde. Pour cela nous avons mesuré le temps que prenait une onde pour parcourir un mètre sur deux cordes, corde A et corde B à l’aide de deux capteurs laser, en tendant plus ou moins la corde, comme vu en Figure 7.

Figure 7 : Montage pour mesurer la vitesse de propagation d'une onde dans une corde
Corde A :
Masse de la corde : $( m_A = 4,6\times10^{-2} \text{kg} $)
Longueur de la corde : $( L_A = 2,08 \text{m} $)
Masse linéique (masse par unité de longueur) : $( \mu_A = \frac{m}{L} = \frac{4,6\times10^{-2}}{2,08} = 2,2\times10^{-2} \text{kg.m}$)-1

Durée Δt (s×10-3) que prend la vibration à parcourir un mètre sur la corde A en fonction de la tension (avec A<B<C<D)
Corde B :
Masse de la corde : $( m_B = 2,1\times10^{-1} \text{kg} $)
Longueur de la corde : $( L_B = 3,07 \text{m} $)
Masse linéique (masse par unité de longueur) : $( \mu_B = \frac{2,1\times10^{-1}}{3,07} = 7,0\times10^{-2} \text{kg.m}$)-1 $( > \mu_A $)

Durée Δt (s×10-3) que prend la vibration à parcourir un mètre sur la corde B en fonction de la tension (avec A'<B')
Dès lors on peut remarquer que plus la tension de la corde est élevée, et plus la propagation se déplace rapidement dans la corde, et plus la corde a une masse linéique élevé, et plus la propagation se déplace lentement.
Le calcul théorique de la vitesse $(v$) d’une onde dans une corde est $( v = \sqrt{\frac{T}{\mu}} $), où $(T$) est la tension de la corde et $(\mu$) sa masse linéique, ce qui est en accord avec nos observations.
Selon une étude menée conjointement par des universités américaines et japonaises, la masse linéique d’une fibre de soie d’araignée est $( \mu_{\text{fibre}} = 1,4 \times 10^{- 8}\text{kg.m}$)-1, même s’il est composé de plusieurs fibres, le fil de soie d’araignée a une masse linéique extrêmement faible. Par ailleurs, la toile est sous tension. Les ondes se déplacent donc extrêmement rapidement dans la toile, ce qui est un des atouts majeurs de la communication vibratoire pour les araignées.
Perception des signaux vibratoires
Les araignées perçoivent très bien les signaux vibratoires, en effet, elles sont dotées de différents organes leur permettant de recevoir des vibrations, notamment les trichobotries et les sensilles en fente, ce sont les organes mécanorécepteurs.
Les trichobotries
Les trichobotries sont des soies sensorielles fines, longues et extrêmement mobiles insérées dans une cupule (une petite structure de la forme d’une coupe), et reliées à un nerf par un dendrite, une courte extension des cellules nerveuses (voir Figure 8).
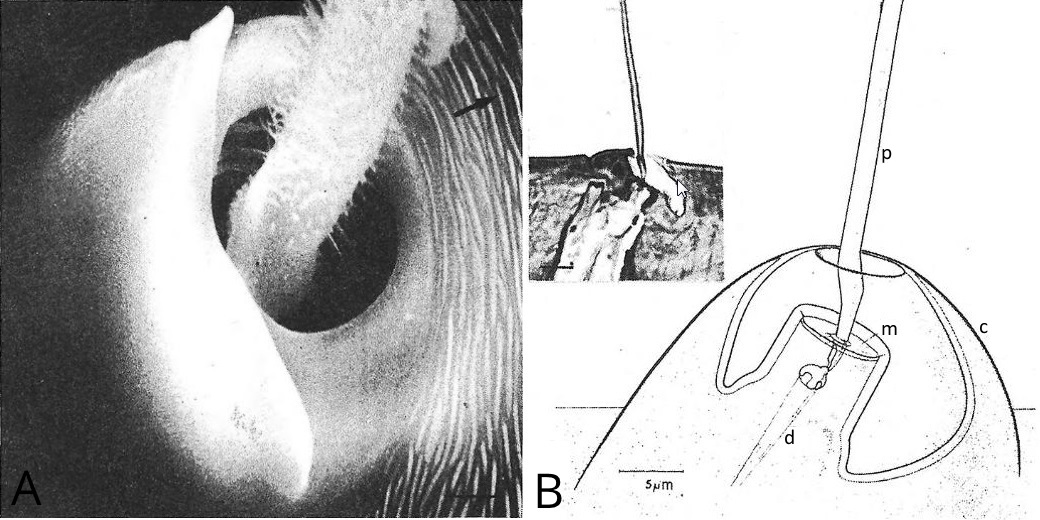
Figure 8 : A : Base d'une trichobotrie, montrant la cupule et la structure plumeuse du poil ; B : Structure d'une trichobotrie (p : poil, c : cupule, m : membrane, d : dentrite)
Les trichobotries sont disposées sur les pattes et les pédipalpes de l’araignée. Ayant une masse très faible, étant extrêmement flexible et étant en contact avec l’air autour de l’araignée, les trichobotries permettent la détection de mouvement dans l’air, mais aussi dans le substrat.
Les sensilles en fente
Les sensilles en fente sont des petits organes mécanorécepteurs qui peuvent détecter la déformation ou la tension. Elles se situent sur l’intégralité de l’exosquelette (aussi appelé cuticule) de l’araignée, et notamment sur les pattes. Leur apparence de fente est à l’origine de leur nom, mais elles ne pénètrent en fait pas l’exosquelette, elles correspondent à un amincissement de la cuticule.
Elles prennent la forme de petites fentes de 1 à 2µm de large et entre 8 et 200µm de long, et peuvent être situées seules ou en groupe (voir Figure 9). Un groupe de sensilles en fente est appelé un organe lyriforme, car, les fentes étant parallèles entre elles, il a la forme d’une lyre, et une sensille en fente seule est appelé sensille en fente simple.
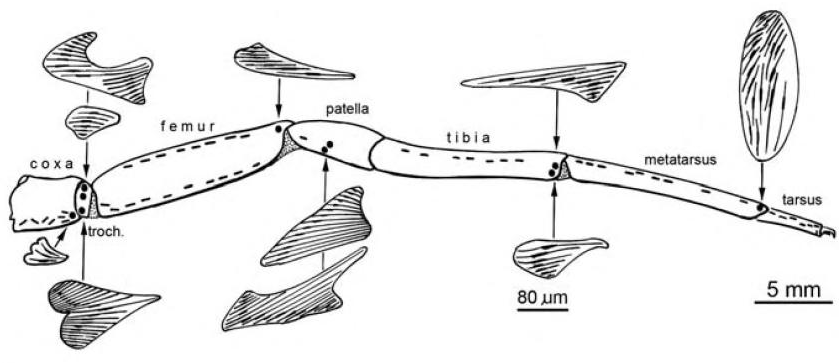
Figure 9 : Distribution des sensilles en fente sur l'arrière de la première patte d'une Cupiennus salei : les traits indiquent des sensilles en fente seule, et les points des organes lyriformes (enlargis pour voir les détails) (d'après Barth et Libera, 1870)
La fente est bordée d’une lèvre cuticulaire et est recouverte d’une membrane cuticulaire. Sous la membrane, une sorte de gouttière s’étend vers le bas et s’élargit en prenant la forme d’une cloche, où se trouvent deux dendrites qui relient la sensille en fente au nerf (voir Figure 10). Même une tension très faible au niveau de la cuticule causera une déformation de la sensille, et la membrane se pliera, déformant l’extrémité d’une des dendrites, provoquant un signal nerveux.

Figure 10 : Schéma d'une sensille simple d'une Cupiennius (d'après Barth, 1971)
C’est en analysant la différence de temps entre l’arrivée de la vibration dans ses différentes pattes que l’araignée peut déterminer la direction de la proie ou de l’intrus.
Utilisation des connaissances en matière de communication vibratoire
Dans la lutte intégrée
Les connaissances en matière de communication vibratoire peuvent être utilisées de différentes manières. En effet, en étant capable de reproduire ou d’imiter des signaux vibratoires, il est possible d’agir sur le comportement des animaux ayant recours à ce type de communication. Ceci est intéressant notamment dans le cadre de la lutte intégrée, définie en Europe par la directive communautaire 91/414/CEE du 15 juillet 1991 ainsi :
« L’application rationnelle d’une combinaison de mesures biologiques, biotechnologiques, chimiques, physiques, culturales ou intéressant la sélection des végétaux dans laquelle l’emploi de produits chimiques phytopharmaceutiques est limité au strict nécessaire pour maintenir la présence des organismes nuisibles en dessous de seuil à partir duquel apparaissent des dommages ou une perte économiquement inacceptables. »
Ainsi, il serait possible de contrôler sans utiliser de pesticides des populations d’insectes nuisibles ayant recours à la communication vibratoire, comme la cicadelle de la vigne (Scaphoideus titanus). Les cicadelles n’utilisent pas de communication chimique à longue portée, mais peuvent transmettre des signaux vibratoires à travers les feuilles de vigne jusqu’à plusieurs mètres de distances. Or, la cicadelle de la vigne est officiellement reconnue comme nuisible car elle est responsable de la propagation de la flavescence dorée, maladie à l’origine de pertes importantes de récoltes de vignes, et son contrôle est donc obligatoire.
Dans son article « Exploitation of Insect Vibrational Signals Reveals a New Method of Pest Management », Anna Eriksson et al. (voir 22 dans la Bibliographie) étudient l’utilisation de vibrations pour perturber la communication entre mâles et femelles Scaphoideus titanus en envoyant des signaux pré-enregistrés que les mâles Scaphoideus titanus utilisent pour empêcher les autres mêles de s’accoupler.

Figure 11 : Nombre de femelles non fécondées trouvées sur les feuilles de vigne (A) sur des plantes en pot, (B) dans un champ de vigne, selon la distance de la source de vibrations (d'après A. Eriksson). Le groupe contrôle est en gris.
Après avoir effectué et analysé des essais, elle a obtenu des résultats concluants que cette méthode de contrôle est très efficace (voir Figure 11) : même lorsque la source de vibration se trouve à 940cm, plus de 80 % des femelles restent non fécondées, alors que dans le cas du groupe contrôle seuls 20 % des femelles restaient non fécondées.
Cette méthode agit donc fortement sur la population, car non seulement elle empêche la fécondation d’une grande partie des femelles, mais elle retarde de plus les fécondations ayant lieu, la femelle est donc moins féconde qu’elle ne l’aurait été. Ainsi, l’utilisation de signaux vibratoires pour perturber la reproduction s’accorde bien à la définition de la lutte intégrée, car en empêchant, ou du moins en retardant la fécondation, elle permet de diminuer la population de nuisible sans l’exterminer, tout en limitant l’utilisation de produits chimiques.
Toutefois, dans une étude plus récente Jernej Polajnar, Anna Eriksson et al. ont mis en évidences l’existence d’effets secondaires causés par la production de signaux vibratoires, notamment la perturbation d’insectes non nuisibles comme les abeilles ou les araignées.
De plus, ils soulignent les limitations techniques de cette méthode : l’atténuation des vibrations est extrêmement rapide dans la plupart des milieux solides. Toutefois certains insectes ont surmonté cette difficulté en produisant des vibrations proches de fréquences résonnantes du substrat, c’est-à-dire les fréquences auxquelles le substrat est sensible, et qui, répétés sous forme périodique, provoquera l’accumulation d’énergie dans le substrat jusqu’à atteindre un régime d’équilibre. Il est donc possible d’imiter les fréquences utilisées par les insectes pour diminuer l’importance du problème d’atténuation.
Par ailleurs, la situation du champ de vignes étudiée par Anna Eriksson était une situation idéale, car les vignes forment une ligne et chaque pied de vigne est relié au suivant, facilitant la transmission de vibrations d’un pied de vigne au suivant. Dès lors, on peut se demander si cette méthode est vraiment applicable.