Introduction
Certaines gammes d’ultrasons peuvent être entendues et utilisées pour communiquer par de nombreux animaux vertébrés terrestres, comme les chiens, certains rongeurs, les chauves-souris ou les dauphins. Ces fréquences peuvent s’étendre de 15 kHz jusqu’à 200 kHz selon les espèces, alors que celles-ci sont audibles par l’homme dans un domaine s’étendant de 20Hz à 20kHz. Les ultrasons dans le monde animal sont étudiés par la bioacoustique. Elle étudie la production, la réception et l’interprétation des sons chez les animaux et humains, mais aussi l’interprétation des sonorités et l’anatomie des corps. Ainsi elle a prouvé que de nombreux animaux utilisent des sons émis non audibles par l’homme. Ces animaux utilisent les ultrasons sous forme de sonar et les utilisent pour se repérer, se nourrir et communiquer. De même, c’est grâce à ces fréquences que certains cétacés et chauves-souris peuvent émettre et réceptionner les ultrasons, ce qui entraine l’exploitation du système d’écholocation par ces espèces.
Une résonance historique
Il y a encore plusieurs dizaines d’année, la production et la perception d’ultrasons étaient considérés possibles uniquement chez les mammifères. En 1973, le scientifique Konishi remarque que les oiseaux n’entendraient pas le son dont la fréquence dépasse 12 kHz, et selon les données disponibles dans les années 1980, l’audition des amphibiens étaient limités à 5 kHz, selon Fay en 1988. Cependant des chercheurs ont ensuite constaté que des amphibiens anoures et un oiseau sont capables de les percevoir. Ce sont la grenouille Odorrana tormota et un passereau chanteur Abroscopus albogularis, ils vivent près de torrents bruyants, et insèrent dans leur chant des harmoniques d’ultrasons. Ainsi, cette grenouille est capable d’émettre et de percevoir des ultrasons, de plus de 100 kHz. C’est la première espèce non mammifère dotée de cette propriété à avoir été découverte. Le mâle pousse des cris semblables à un chant d’oiseau et possède une anatomie de l’oreille inhabituelle, avec notamment un tympan concave, qui contient un canal auditif et des osselets très légers, donc très sensibles.
L’écholocation
L’écholocalisation consiste à envoyer des sons et à écouter leur écho pour localiser et identifier les éléments d’un environnement. Certaines espèces peuvent émettre des ultrasons comme les chauves-souris. Elles émettent des ultrasons qui se répercutent sur les objets environnants, ce qui leur permet ainsi de percevoir leur environnement. Ainsi la chauve-souris se sert de l’écholocation pour se repérer dans l’obscurité et pour repérer sa proie. Elle émet des impulsions à haute fréquence. Le son récupéré sur la trajectoire de la chauve-souris revient vers celle-ci. L’écholocation permet à la chauve-souris de déterminer la proximité de l’insecte. Grâce à la réponse continue des impulsions réfléchies, la chauve-souris se dirige automatiquement vers la proie pour la capturer.
L’écholocation a trois principaux inconvénients. D’une part, elle nécessite la réception et le traitement d’échos très faibles ; cela implique un appareil auditif très perfectionné (d’où les oreilles très grandes des chauves-souris) et un appareil neurologique de traitement très fin. Pour qu’il puisse entendre un écho, il est nécessaire à l’émetteur de produire des cris très forts, d’où des fréquences élevées.
D’autre part, la cible peut percevoir les sons émis pour la repérer et réagir en s’échappant.
Enfin, chez les animaux vivant en groupe (chauve-souris et dauphins notamment), l’émission et la réception de leurs signaux peuvent être brouillés par ceux de leurs congénères.
L’exemple des dauphins
L’écholocation des dauphins est la capacité de ces deniers à repérer et situer les aspects importants de leur environnement, leurs congénères ou les proies. Elle se base sur la propagation des ondes dans l’eau.
Le dauphin est capable d’émettre différents types de son, de fréquences variables, certains servant à communiquer, d’autres à se repérer dans l’espace. Le système d’émission chez le dauphin est bien plus complexe que chez l’homme. L’homme n’est en effet capable de produire que des sons qui luis sont audibles, c’est-à-dire entre 20 et 20 000 Hz.
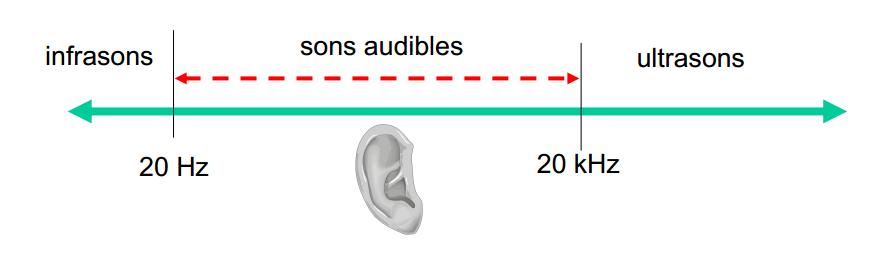
Schéma des fréquences des ultrasons
On peut séparer les ondes acoustiques émises par les dauphins en deux grands groupes : les sifflements, utilisés pour communiquer, et les clics servant à l’écholocation. Le langage des dauphins est mal connu et semble complexe. Tout au plus peut-on dire qu’il présente de grandes variations en fonction du groupe de dauphins étudié et des individus. Les sifflements sont bien localisés en fréquence et se situent plutôt dans les ultrasons (< 25 kHz).
Les sifflements sont des sons caractéristiques d’une espèce ou d’un individu. Ce sont des faisceaux couvrant une bande étroite de basse fréquence, et utilisés pour la communication entre espèces.
Les clics d’écholocation sont des signaux très brefs (quelques dizaines de microsecondes) et donc répartis sur une large bande spectrale. Par exemple la largeur de bande à 3 dB renvoie à une cinquantaine de kHz. Plus le dauphin se rapproche de sa proie, plus le train de clics est rapide. La résolution maximale que peut traiter le cerveau du dauphin est de 600 clics par secondes.
Ces clics peuvent être représentés selon deux types. Le premier est sous la forme d’un faisceau large de basse fréquence, d’émission lente et de longue portée. Ils sont utilisés pour créer des images de son environnement. Le second type des clics d’écholocation est représenté par un faisceau étroit à courte portée, occupant une large bande de haute fréquence. Ils servent à analyser une potentielle proie ou un objet rencontré.
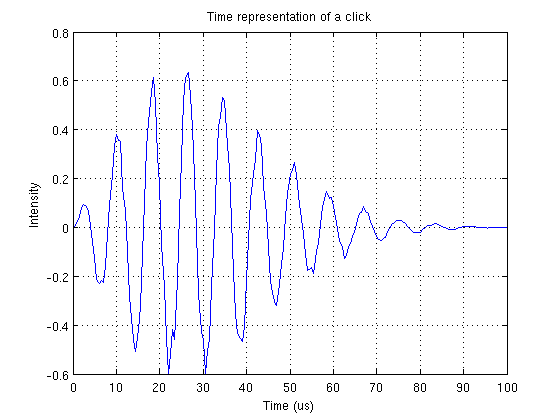
Représentaion temporelle d'un clic 1
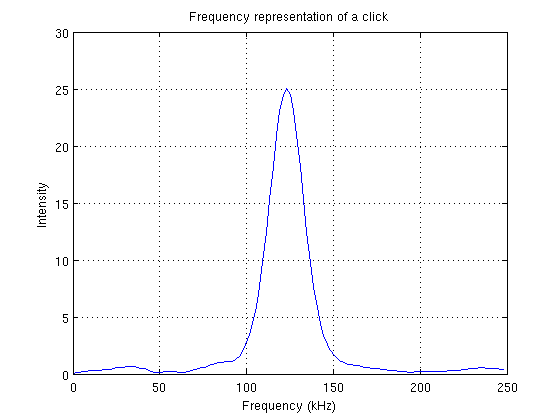
Représentation fréquentielle d'un clic 1
L’émetteur
Le dauphin ne possède pas de cordes vocales mais trois pairs de sacs aériens, de forme différente, situés sous l’évent, de part et d’autre du conduit nasal. Ces sacs servent de réserves d’air. Le dauphin arrive à produire des sons en faisant passer l’air d’un sac à l’autre par des ouvertures dont il règle le diamètre, grâce à un ensemble de muscles et de nerf. Pour pouvoir orienter de manière précise ses faisceaux sonores le dauphin est doté d’un organe, le melon. Les ondes sonores créées vont se réfléchir sur la paroi osseuse frontale concave du crâne de l’animal pour ensuite se concentrer sur une masse graisseuse qui est le melon. Le dauphin peut ainsi orienter l’émission de ses ondes ultrasonores où il le souhaite.
Le larynx est aussi utilisé pour l’émission sonore. Le dauphin peut en effet expulser de l’air par ses poumons et cet air fait vibrer des muscles puissants du conduit respiratoire. Ces muscles vont alors transmettre ces vibrations à des os dont ceux des maxillaires.
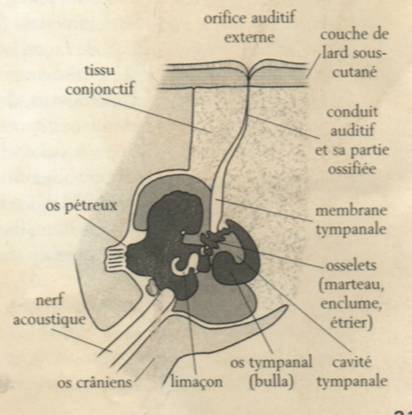
Anatomie du système d'émission des ultra-sons
Le récepteur
Le système de réception du dauphin est complexe. Il repose sur deux récepteurs qui sont les oreilles et le maxillaire inférieur
Les sons sont réceptionnés au niveau de l’orifice auditif externe. Ils passent par le conduit auditif et arrivent dans l’oreille interne.
Ils sont ensuite transmis au cerveau par l’influx nerveux d’un nerf acoustique. Les centres acoustiques du cerveau dont le but est d’analyser les messages sonores sont très développés chez le dauphin. Tout le système auditif de cet animal est protégé par des masses de mucus qui arrêtent les vibrations parasites qui pourrait provenir de l’environnent.
Les ondes acoustiques peuvent aussi être reçues par le maxillaire inférieur, les sons s’y propagent mieux. Les ondes sonores sont donc reçues par l’extrémité du bec de l’animal, elles se propagent dans un corps graisseux au niveau du maxillaire et sont transmises à l’oreille interne au niveau de l’articulation de la mâchoire. Ensuite, tout ce passe comme ce qui a été expliqué précédemment.
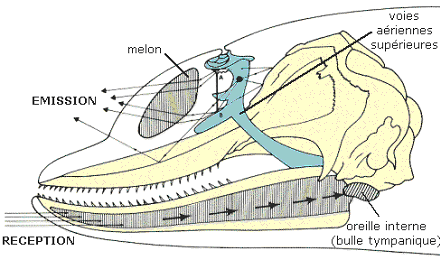
Profil d'une tête de dauphin
L’effet Doppler
Pour recevoir les signaux réfléchis par les cibles, le dauphin exploite des tissus adipeux situés sous sa mâchoire, qui remontent jusqu’à son oreille interne. Le son est donc transmis à l’oreille interne, puis au cerveau, qui l’analyse. Le dauphin peut alors déterminer la distance de la cible, sa taille, ainsi que sa vitesse en mesurant la différence de fréquence en exploitant l’effet Doppler. C’est à dire le décalage de fréquence d’une onde entre la mesure à l’émission et la mesure à la réception lorsque la distance entre l’émetteur et le récepteur varie au cours du temps. Le dauphin peut aussi sonder sous les sédiments, étant donné que le son se propage sous le sable.
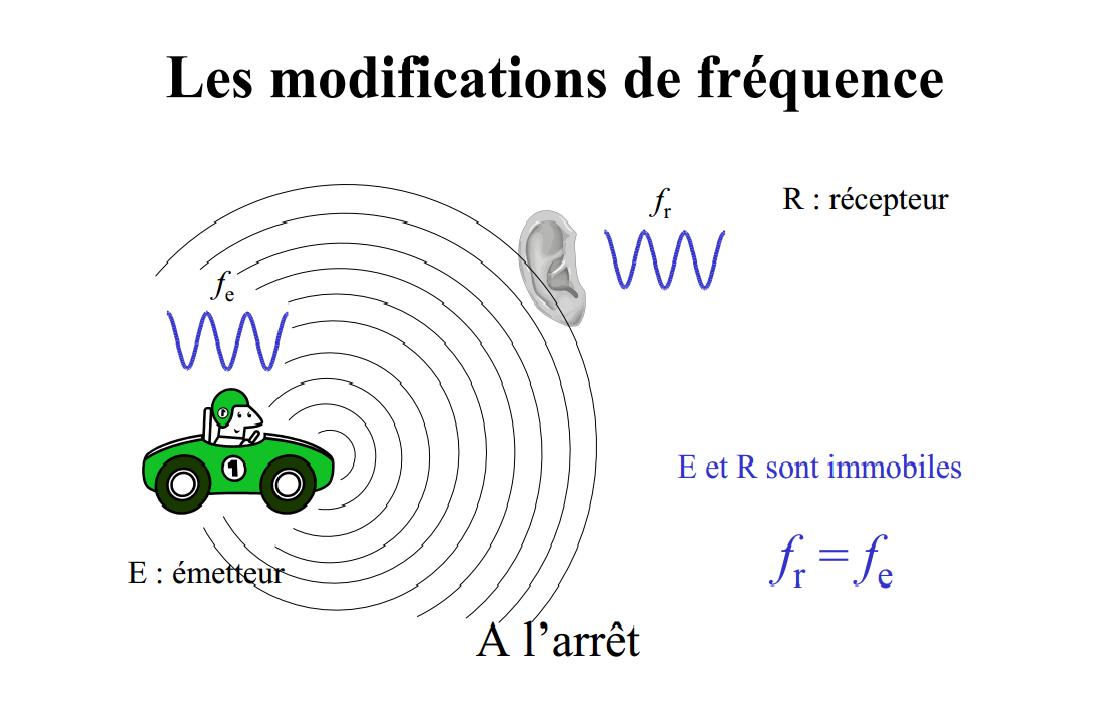
L’effet Doppler est le décalage de fréquence d’une onde acoustique ou électromagnétique entre la mesure à l’émission et la mesure à la réception lorsque la distance entre l’émetteur et le récepteur varie au cours du temps elle renseigne aussi sur la vitesse de la cible par rapport à l’émetteur.
Mise en image de l'effet Doppler : « à l'arrêt »
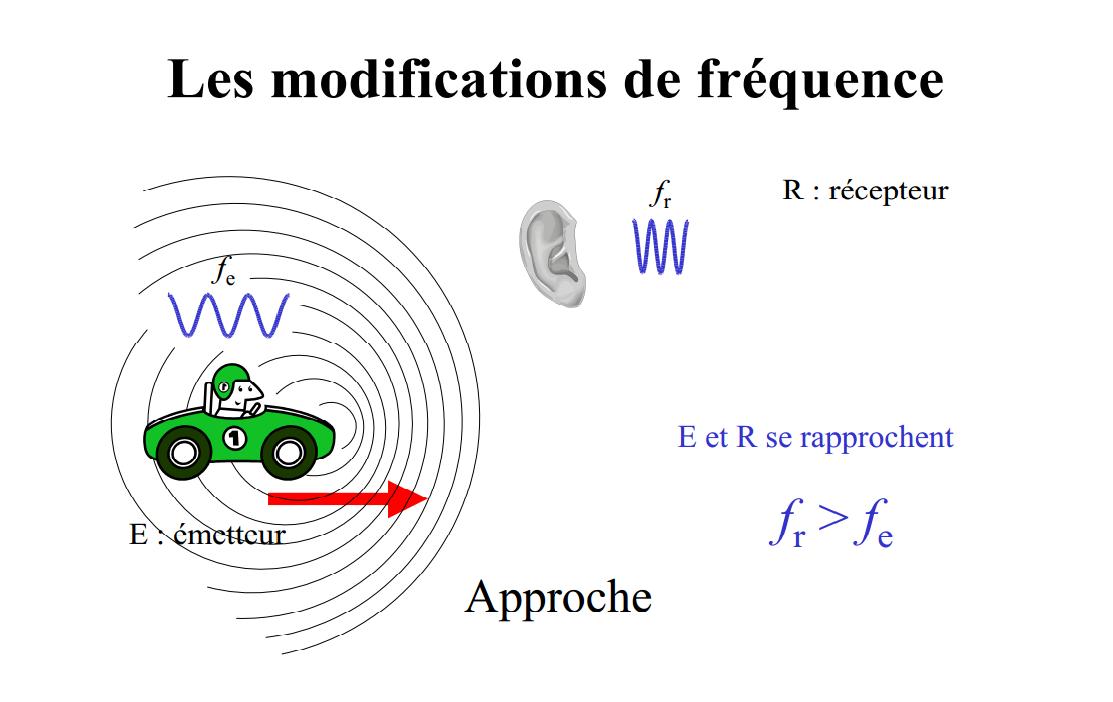
La la longueur d’onde du signal perçu dépend de la vitesse relative entre la source et le récepteur.
Si un observateur en mouvement cherche à mesurer cette longueur, il lui trouve une valeur différente, plus élevée s’il se déplace dans le sens de l’onde, plus courte s’il se déplace en sens contraire.
Mise en image de l'effet Doppler : « en approche »
L'effet Doppler
Si la vitesse $( V_{r} $) (vitesse de l’objet mobile perçue par l’observateur) est petite devant $(c$) (vitesse de l’onde perçue), en termes de longueur d’onde $( \lambda = c \times T $), l’observateur perçoit un rayonnement de longueur d’onde :
$[ \lambda' = \lambda \times (1 + \frac{V_{r}}{c}) $]
Soit un rayonnement décalé de la quantité
$[ \Delta\lambda = \lambda' - \lambda $]
Tel que
$[ \Delta\lambda = \frac{V_{r}}{c}\times\lambda $]
Ce calcul montre que le décalage relatif de la longueur d’onde est proportionnel au rapport de la vitesse de la source par rapport à l’observateur à la vitesse de la lumière.
La prestine
Chez plusieurs espèces douées de compétence d’écholocation par émission et réception d’ultrasons, dans l’eau ou dans l’air ; certains cétacés et chauve-souris ont recours à la prestine, protéine impliquée dans l’écholocation.
Des chercheurs se sont concentrés sur cette protéine présente chez tous les mammifères. Elle est située sur les cellules cillées externes de la cochlée, dans l’oreille interne, cette protéine joue un rôle d’amplificateur des ondes sonores (en particulier des hautes fréquences comme les ultrasons), grâce aux vibrations qu’elle provoque. Cette protéine est similaire chez les différents groupes de chauves-souris qui utilisent l’écholocation.
L’équipe sino-britannique de Stephen Rossiter (University of London, GB) et l’équipe sino-américaine de Jianzhi Zhang (University of Michigan, E-U) ont comparé la prestine des chauves-souris et celle de différents cétacés.
En construisant des arbres phylogénétiques uniquement basés sur l’évolution de la prestine (et non sur les autres caractéristiques de ces mammifères), les deux équipes ont abouti à des rapprochements surprenants. Les chauves-souris et dauphins doués pour l’écholocation devenaient cousins, formant un groupe évolutif cohérent.
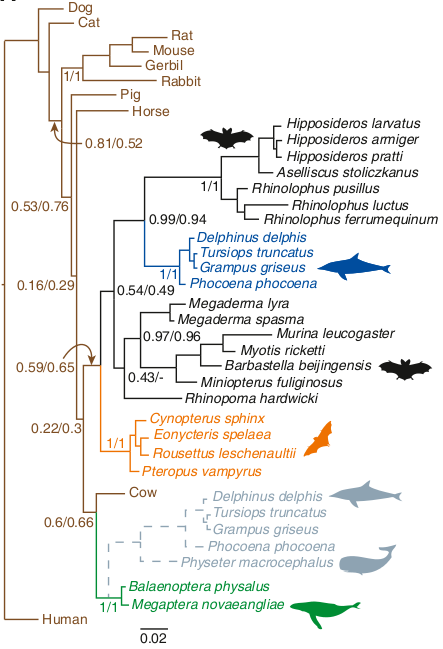
Arbres phylogénétiques marquant l’évolution de la prestine
Les dauphins (en bleu) au milieu des chauves-souris (en noir) et donc éloignées des baleines dépourvues d’écholocation (en bas en vert).
Ces chercheurs avaient analysés la suite de nucléotides composant le gène de la prestine chez cinq espèces d’odontocètes, le cachalot et quatre espèces de dauphin, ainsi que chez des baleines incapables d’écholocalisation. Puis ils avaient comparé les séquences obtenues à celles de 18 espèces de chauves-souris. Ils avaient alors mis en évidence une ressemblance entre les séquences des odontocètes à sonar, le cachalot mis à part, et celles des chauves-souris.
Donc cela signifie que la version de la protéine codée par le gène Prestin chez les dauphins est plus proche de la version de la protéine chez les chauves-souris douées d’écholocalisation que de la version de la protéine chez les baleines dépourvues d’écholocalisation. Mais comment est-ce possible ?
Au cours de l’évolution, des mutations se sont accumulées dans l’ADN du gène Prestin. Lorsqu’une mutation survient dans un gène, elle modifie le codon en question, ce qui peut avoir pour conséquence de changer l’acide aminé.
Si l’acide aminé n’est pas modifié, il n’y a pas de conséquence sur le phénotype. Par contre, si l’acide aminé est modifié, la protéine a de grandes chances de fonctionner différemment. Si elle fonctionne mieux, l’organisme se reproduira en moyenne plus que les autres organismes. Grâce à l’action de la sélection naturelle, une mutation positive peut être de plus en plus présente dans la population au cours des générations.
Dans le cas des dauphins et des chauves-souris, les chercheurs cités ci-dessus ont montré que, par la sélection naturelle, les différences entre leurs séquences d’ADN du gène Prestin correspondent à des changements vers les mêmes acides aminés. A l’inverse, les différences entre les séquences d’ADN des dauphins et des baleines dépourvues d’écholocalisation correspondent à des changements d’acides aminés différents.
Cela signifie que la prestine joue un rôle important dans l’écholocalisation, que celle-ci ait été acquise dans l’eau ou dans l’air. Les mutations du gène de la prestine doivent avoir été suffisamment fréquentes pour que les individus dotés par hasard de la bonne séquence de la protéine, celle qui est associée à la meilleure sensibilité aux ultrasons, soient sélectionnés par leur environnement.
Utilisation de ces connaissances par l’homme
United States Navy Marine Mammal Program
En 1942, la Suède a eu l’idée d’utiliser des phoques pour faire la chasse aux sous-marins allemands. Ces phoques dressés, équipés d’une mine sur leur dos, devaient nager sous les coques, puis le moment venu, la mine explosait avec le corps de l’animal.
Cette nouvelle tactique de guerre a ouvert la voie aux expériences militaires. L’US Navy utilise des otaries et des dauphins depuis 1960.

Le programme de mammifères marins de la marine américaine, en anglais U.S. Navy Marine Mammal Program (NMMP), est un programme dirigé par la Marine américaine qui étudie l’emploi militaire de mammifères marins (le Grand dauphin qui émet et reçoit en un millième de seconde, des fréquences variant entre 220 et 250 000 hertz et l’otarie de Californie) et les entraîne à des tâches tels que la protection de navires et de ports, le repérage et le dégagement de mines, ainsi que la récupération d’objets. Le programme est basé à San Diego en Californie où les animaux sont entraînés. Les animaux du NMMP ont été déployés en zones de combat, notamment pendant la Guerre du Viêt Nam et la Guerre d’Irak. Cette dernière a permis aux américains d’ouvrir le golfe Arabique au marché international après la fin du conflit, en particulier grâce a ces animaux.La Navy a recours à certaines équipes humains-crétacés. Les équipes MK 4, 7 et 8 utilisent des dauphins; MK 5 utilise des otaries, et MK 6 utilise à la fois dauphins et otaries. Ces équipes peuvent être déployées partout sur le globe en 72 heures, vers des zones de conflits.

Soldat dauphin
Les ondes dans le milieu liquide
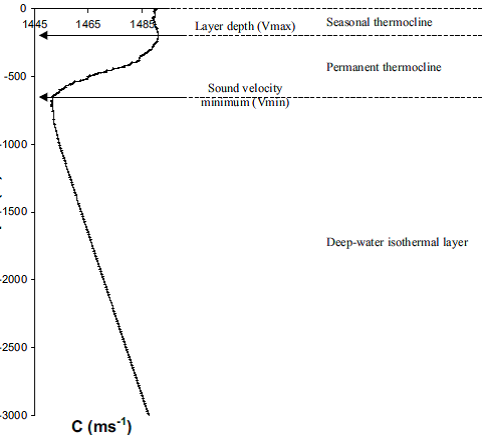
Niveaux de profondeur dans l'eau
Trois niveaux de profondeur sont visibles : la zone proche de la surface (“seasonal thermocline”) dont la température dépend de la saison de l’année, la zone intermédiaire thermocline (“permanent thermocline”) puis la zone des eaux froides profondes (“deep-water isothermal layer”).
Passée la valeur de la vitesse minimale (“sound velocity minimum” : Vmin), la profondeur a donc une influence positive sur la célérité.
Nous pouvons remarquer que la vitesse de propagation des ondes sonores dans l’eau n’est pas constante. Elle est maximale dans les eaux de surface puis décroit dans la zone intermédiaire jusqu’à sa vitesse minimale pour augmenter de nouveau dans les eaux profondes.
Ces données peuvent justifier ceci grâce à ces données :
-
Dans l’air, le signal reçu a un retard de 600 µs sur le signal émis.
-
Dans l’eau, le signal reçu a un retard de 150 µs sur le signal émis.
Ainsi les ultrasons se déplacent plus rapidement dans l’eau que dans l’air, à raison de 300m/s dans l’air et de 1500m/s dans un milieu liquide comme l’eau.
L’échographie
L’échographie est une méthode d’exploration du corps reposant sur l’utilisation des ultrasons. Grâce à la sonde qui émet et reçoit les ultrasons, l’échographie permet une représentation interne du corps. Elle est principalement utilisée en médecine. Nous allons désormais évoquer l’examen “Doppler vasculaire”.
Un examen “Doppler vasculaire” se pratique au moyen d’une sonde Doppler qui émet ses ultrasons à une fréquence précise en direction de l’artère à étudier. Ces ultrasons percutent les hématies en mouvement, or la taille des hématies est inférieure à la longueur d’onde des ultrasons utilisés. Il se produit donc un phénomène de diffusion.
L’énergie rediffusée par les hématies revient vers la sonde qui va mesurer la fréquence de ces ondes. Cette nouvelle fréquence sera différente de celle des ultrasons émis en raison de l’effet Doppler qui intervient deux fois : d’abord entre la sonde (l’émetteur fixe) et les hématies (récepteurs mobiles) ; puis entre ces hématies, qui deviennent les “émettrices” mobiles et la sonde (récepteur fixe).
La différence entre la fréquence du signal émis et celle du signal reçu par la sonde suit la formule:
$[ \delta F = 2F\left(\frac{V}{C}\right) \cos \theta $]
Avec
$( \delta F $) : différence entre la fréquence de l’onde émise et celle de l’onde reçue
$(F$) : fréquence de l’onde émise
$(V$) : vitesse du sang
$(C$) : célérité des ultrasons dans les tissus mous (1540 m/s)
$(\cos \theta$) : cosinus de l’angle que forme la direction du faisceau ultrasonore et la direction de l’écoulement sanguin.